Zakład Żywienia Zwierząt

Kierownik Zakładu
dr hab. Barbara Kowalik, profesor instytutu
🖂 b.kowalik@ifzz.pl
Pracownicy naukowi
prof. dr hab. Marian Czauderna, 🖂 m.czauderna@ifzz.pl
dr hab. Grzegorz Bełżecki, profesor instytutu, 🖂 g.belzecki@ifzz.pl
dr hab. inż. Małgorzata Białek, profesor instytutu, 🖂 m.bialek@ifzz.pl
dr hab. inż. Paweł Konieczka, prof. UWM, 🖂 p.konieczka@ifzz.pl
dr hab. Paweł Kowalczyk, 🖂 p.kowalczyk@ifzz.pl
dr hab. Renata Miltko, profesor instytutu, 🖂 r.miltko@ifzz.pl
dr hab. Grzegorz Skiba, profesor instytutu, 🖂 g.skiba@ifzz.pl
dr hab. Ewa Święch, profesor instytutu, 🖂 e.swiech@ifzz.pl
dr hab. Anna Tuśnio, profesor instytutu, 🖂 a.tusnio@ifzz.pl
dr inż. Marcin Barszcz, 🖂 m.barszcz@ifzz.pl
dr Małgorzata Majewska, 🖂 m.majewska@ifzz.pl
dr inż. Monika Sobol, 🖂 m.sobol@ifzz.pl
mgr Kamil Gawin, 🖂 k.gawin@ifzz.pl
mgr Misza Kinsner, 🖂 m.kinsner@ifzz.pl
mgr Adrianna Konopka, 🖂 a.konopka@ifzz.pl
mgr Wiktoria Wojtak, 🖂 w.wojtak@ifzz.pl
Pracownik badawczo-techniczny
dr hab. Agnieszka Białek 🖂 a.bialek@ifzz.pl
Pracownicy inżynieryjni
inż. Natalia Frączek, 🖂 n.fraczek@ifzz.pl
mgr Aneta Kędzierska, 🖂 a.kedzierska@ifzz.pl
mgr inż. Urszula Wolska-Świętlicka, 🖂 u.wolska-swietlicka@ifzz.pl
Pracownicy techniczni
Malwina Krakowiak, 🖂 m.krakowiak@ifzz.pl

Aktualna tematyka badawcza
W Zakładzie prowadzone są badania głównie na: drobiu, trzodzie chlewnej, przeżuwaczach oraz na zwierzętach laboratoryjnych i mikroorganizmach zasiedlających przedżołądki zwierząt przeżuwających.
- Identyfikacja i charakterystyka drobnoustrojów zasiedlających przewód pokarmowy ptaków i ssaków.
- Izolacja i hodowla mikroorganizmów w warunkach in vitro.
- Określenie zmian jakościowych i ilościowych symbiotycznej mikroflory i mikrofauny pod wpływem bioaktywnych składników pokarmu.
- Rola orzęsków żwaczowych w procesach metanogenzy i gospodarki lipidowej.
- Izolacja i charakterystyka enzymów pochodzenia mikrobiologicznego katalizujących trawienie węglowodanów zapasowych i strukturalnych (pochodzenia roślinnego i mikrobiologicznego) w żwaczu.
- Określenie wpływu dodatków paszowych na wskaźniki procesów trawiennych oraz wyniki produkcyjne i jakość mięsa oraz na rozkład składników pokarmowych pasz metodą in sacco.
- Wykorzystanie metod in vitro w badaniach strawności składników pokarmowych i aktywności flory bakteryjnej.
- Wykorzystanie krajowych źródeł białka w żywieniu ptaków i ssaków.
- Wpływ procesów technologicznych na wartość odżywczą nasion roślin bobowatych.
- Badania nad wpływem włókna pokarmowego na fizjologię przewodu pokarmowego.
- Rola czynników żywieniowych na właściwości kości rosnących zwierząt.
- Wpływ związków bioaktywnych na układ odpornościowy przewodu pokarmowego zwierząt monogastrycznych i przeżuwających.
- Wpływ diety w zapobieganiu występowania stresu oksydacyjnego w organizmach zwierząt.
- Wpływ olejów oraz antyutleniaczy na profil kwasów tłuszczowych, aminokwasów, status antyoksydacyjny i zawartość cholesterolu w wybranych tkankach i narządach zwierząt.
- Wpływ dodatku izomerów CLA do diety matek z chorobą nowotworową na stan zdrowia potomstwa – badania na modelu szczura (programowanie żywieniowe).

Najważniejsze osiągnięcia
- Identyfikacja i charakterystyka nowego gatunku bakterii Treponema zioleckii oraz opracowanie metody określania biomasy bakterii na podstawie zawartości mureiny (Piknova i wsp., 2008. FEMS Microbiol. Lett. 289, 166-172; Bełżecki i wsp., 2012. J. Anim. Feed Sci. 21, 624-634).
- Wykazano, że orzęski żwaczowe mogą być wykorzystywane, jako potencjalne źródło wodoru w ogniwach paliwowych służących do pozyskiwania energii elektrycznej oraz, że dysponują one kompletem enzymów zdolnych do rozkładu węglowodanów, w tym chityny (Piela i wsp., 2010.
J. Microbiol. Biotechnol. 20, 1092-1100; Miltko i wsp., 2015. J. Anim. Feed Sci. 24, 203-207; Miltko i wsp., 2016. Arch Anim Nutr. 70, 425-440). - Wykazano, że dodatek kory dębu (Quercus sp.) do dawek pokarmowych dla owiec zwiększa ogólną liczebność pierwotniaków w żwaczu oraz, że stymuluje aktywność zewnątrz-wydzielniczą trzustki. Ponadto wykazano, że liście borówki (Vaccinium vitis idaea L.) stosowane w dawkach pokarmowych dla jagniąt poprawiają jakość mięsa (Majewska i wsp., 2021. Anim. Biosci. 34, 1146-1156; Majewska i wsp., 2017. J. Anim. Feed Sci. 26, 354-358; Majewska i Kowalik, 2020. Eur. J. Lipid Sci. Tech. 122, 1900273).
- Wykazano, że dodatek olejów roślinnych o wysokiej zawartości kwasu α-linolenowego (olej lniany) i oleinowego (olej rzepakowy) w dawkach pokarmowych dla owiec istotnie zmniejsza rozkład celulozy. Stwierdzono również, że olej rzepakowy negatywnie wpływa na aktywność pektynolityczną w żwaczu owiec. Ponadto, wykazano, że zastosowane oleje zwiększają aktywność lipazy trzustkowej oraz, że olej lniany bardziej niż olej rzepakowy poprawia jakość jagnięciny (Majewska i wsp., 2020. Livest. Sci. 240, 104175; Kowalik i wsp., 2018. J. Anim. Physiol. Anim. Nutr. 102, 1194-1198; Miltko i wsp., 2019. Asian-Austral. J. Anim. Sci. 32, 767-775).
- Wykazano, że kwas karnozynowy i selenian (VI) dodane do dawek pokarmowych zmniejszają biosyntezę CH4 i CO2 w żwaczu owiec oraz, że poprawiają wykorzystanie paszy przez jagnięta. Ponadto, stwierdzono, że zastosowanie selenianu (IV) zmniejsza biouwodornienie nienasyconych kwasów tłuszczowych w żwaczu owiec (Białek i Czauderna, 2019. Livest. Sci. 226, 122-132; Miltko i wsp., 2016. Acta Vet.-Beograd 66, 373-391; Białek i wsp., 2020. Livest. Sci. 241, 104249).
- Wykazano, że dodanie do diety jagniąt oleju rybnego w połączeniu z kwasem karnozynowym i organicznym selenem zwiększa gęstość oraz poprawia właściwości biomechaniczne kości (Skiba i wsp., 2021. Animals 11, 230).
- Wykazano, że fruktany typu inulinowego mogą przyczyniać się do poprawy dobrostanu prosiąt przez zwiększanie populacji bakterii z rodzaju Bifidobacterium, działanie antyoksyacyjne oraz korzystne zmiany ekspresji białek w błonie śluzowej jelita grubego świń. Wskazuje to na korzystny wpływ na funkcjonowanie bariery jelitowej, ochronę przed endotoksynami i wolnymi rodnikami, a tym samym na możliwość stosowania inuliny w dietoprofilaktyce chorób jelita grubego (Barszcz i wsp., 2016. Arch. Anim. Nutr. 70, 278-292; Barszcz i wsp., 2018. Anim. Prod. Sci. 58, 1109-1118; Herosimczyk i wsp., 2020. Animal 14, 1647-1658).
- Wykazano, że dodatek glicynianu cynku do paszy dla prosiąt sprzyja rozwojowi populacji Clostridium herbivorans, a dodatek włókna ziemniaczanego zwiększa aktywność bakteryjnej β-glukozydazy w jelicie grubym, co przyczynia się do zwiększenia strawności włókna pokarmowego (Barszcz i wsp., 2019. Livest. Sci. 227, 37-43; Barszcz i wsp., 2021. Livest. Sci., 245, 104429).
- Wykazano, że zarówno wysoki poziom łubinu wąskolistnego w paszy, jak i proces mikronizacji mogą mieć negatywny wpływ na skład i aktywność mikroflory jelita grubego prosiąt. Ponadto stwierdzono, że wysoki poziom łubinu w mieszankach może mieć niekorzystny wpływ na przyrost masy ciała prosiąt (Tuśnio i wsp., 2020. Livest. Sci. 240, 104137).
- Wykazano, że interakcje pomiędzy źródłem białka a źródłem węglowodanów
w paszy, wpływają na aktywność flory bakteryjnej jelita grubego świń. Modyfikacja fermentacji proteolitycznej za pomocą węglowodanów złożonych jest możliwa, lecz kierunek tych zmian zależy od rodzaju węglowodanów. Surowa skrobia ziemniaczana pogłębia negatywny wpływ białka gorzej trawionego w jelicie cienkim, natomiast pektyna i celuloza zmniejszają intensywność bakteryjnej proteolizy (Taciak i wsp., 2017. Arch. Anim. Nutr. 71, 192-209). - Wykazano, że dodatek inuliny redukuje negatywny wpływ wysokotłuszczowej diety bogatej w nasycone kwasy tłuszczowe na właściwości kości rosnących świń oraz cechy prozdrowotne i technologiczne mięsa wieprzowego oraz określono zależności między proporcją n-6/n-3 PUFA w paszy i ciele a właściwościami kości rosnących świń (Sobol i wsp., 2018. Br. J. Nutr., 119: 1111-1118; Przybylski i wsp., 2019. J. Anim. Physiol. Anim. Nutr., 103, 593-602; Sobol i wsp., 2019. Br. J. Nutr., 121, 508–518).
- Wykazano, że niska proporcja kwasów PUFA n-6/n-3 oraz zwiększona dawka witaminy E w diecie kurcząt powodują uszkodzenia DNA komórek nabłonka jelit oraz negatywnie oddziałuje na jego mikrostrukturę. Ponadto, wykazano, że zwiększenie w czasie odchowu zawartości witaminy E w paszy powoduje oporność kurcząt brojlerów z indukowanym stanem zapalnym na działanie kwasu acetylosalicylowego. Przeprowadzona po raz pierwszy u drobiu analiza zużycia tlenu przez mitochondria płytek krwi udowodniła, że podanie kwasu acetylosalicylowego nie wpływa negatywnie na funkcję mitochondriów, a wręcz przeciwnie, może przeciwdziałać negatywnym skutkom stanu zapalnego (Konieczka i wsp., 2018. Poult. Sci. J., 97, 149-158; Konieczka i wsp., 2019. Vet. Res. 50, ID 65).
- Wykazano, że nano selen (Nano-Se) oraz ekstrakt z konopi siewnych (Cannabis sativa L.) stosowane w żywieniu kurcząt brojlerów mogą wspierać funkcje bariery jelitowej u ptaków narażonych na infekcję bakteriami C. perfringens, które są odpowiedzialne za wywoływanie martwiczego zapalenia jelit u drobiu (Konieczka i wsp., 2020. Vet. Res. 51, ID 141).
- Wykazano, że w sercach szczurów z indukowanym nowotworem sutka, izomery CLA zawarte w dietach, wpływają hamująco na peroksydację wielonienasyconych kwasów tłuszczowych oraz obniżają poziom aterogennych nasyconych kwasów tłuszczowych i zmniejszają utlenianie cholesterolu (Białek i wsp., 2019, Nutrients 11, 2032; Białek i wsp., 2020, Animals 10, 464).
- Wykazano, że olej z nasion granatowca właściwego powoduje zmiany w składzie i zawartości kwasów tłuszczowych, obniża aktywność desaturaz i ogranicza powstawanie produktów oksydacji lipidów w wątrobie szczurów (Białek i wsp., 2017. Prostagland. Other Lipid Mediat. 131, 9-16).
- Wykazano, że pochodne kumaryny, 1.2 alfa-aminoamidy, diaryloetanole oraz δ-lactony działają toksycznie na komórki bakteryjne E. coli K12, R1-R4 występujące w przewodzie pokarmowym. Związki te, indukują w komórkach bakterii patogennych stres oksydacyjny i hamują ich replikację. Stwierdzono również, że badane związki charakteryzują się większą skutecznością i selektywnością względem określonych szczepów bakteryjnych E.coli, jednocześnie wykazując większą stabilność biologiczną oraz toksyczność dla organizmu człowieka w porównaniu do tradycyjnie stosowanych antybiotyków (Kowalczyk i wsp., 2020. Materials 13, 2499; Kowalczyk i wsp., 2020. Materials 13, 5169; Kowalczyk i wsp., 2021. Materials 14, 1025; Kowalczyk i wsp., 2021. Materials 14, 2956).
Wybrane publikacje
- Barszcz M., Tuśnio A., Taciak M., 2024. Poultry nutrition. Physical Sciences Reviews. 9, 2, 611-650, https://doi.org/10.1515/psr-2021-0122
MNiSW 20 pkt. - Bełdowska A., Pietrzak E., Biesek J., Barszcz M., Tuśnio A., Konopka A., Gawin K., Dunisławska A. 2024. The effect of sodium butyrate administered in ovo on the health status and intestinal response in broiler chicken. Poultry Science 103, 104108, https://doi.org/10.1016/j.psj.2024.104108
MNiSW 140 pkt. IF 3,8 Q1 - Bełdowska A., Siwek M., Biesek J., Barszcz M., Tuśnio A., Gawin K., Dunisławska A. 2024. Impact of in ovo administration of xylo- and mannooligosaccharides on broiler chicken gut health. Poultry Science 103, 104261, https://doi.org/10.1016/j.psj.2024.104261 MNiSW 140 IF 3,8 Q1
- Białek M., Białek A., Wojtak W., Czauderna M. 2024. Organic and inorganic selenium compounds affected lipidomic profile of spleen of lambs fed with diets enriched in carnosic acid and fish oil. Animals14, 1, 133, https://doi.org/10.3390/ani14010133 MNiSW 100 IF3,0 Q1
- Białek M., Lepionka T., Wojtak W., Ruszczyńska A., Bulska E., Czauderna M. and Białek A., 2024. Splenic elemental composition of breast cancer-suffering rats supplemented with pomegranate seed oil and bitter melon extract. Molecules29, 1942, https://doi.org/10.3390/molecules29091942 MNiSW 140 IF 4,2 Q1
- Bień D., Michalczuk M., Jóźwik A., Matuszewski A. Konieczka P. 2024. Effects of cannabis sativa extract on growth performance, meat physicochemical properties, and oxidative status in chickens challenged with Clostridium perfringens and lipopolysaccharide. Animal Science Papers and Reports 42, 1: 81-108, https://doi.org/10.2478/aspr-2023-0024 MNiSW 100 IF 0,9 Q3
- Chodkowska K.A., Barszcz M., Tuśnio A. 2024. MicroRNA expression and oxidative stress markers in pectoral muscle of broiler chickens fed diets supplemented with phytobiotics composition. Scientific Reports14, 4413, https://doi.org/10.1038/s41598-024-54915-y
MNiSW 140 pkt. IF 3,8 Q1 - Czauderna M., Taubner T., Wojtak W. 2024. Comparative study of gas and liquid chromatography methods for the determination of underivatised neutral and acidic cannabinoids and cholesterol. Molecules 29, 2165, https://doi.org/10.3390/molec ules29102165 MNiSW 140 pkt. IF 4,2 Q1
- Czauderna M., Wojtak W., Białek M., Białek A. 2024. Optimization of high-efficient pre-column sample treatments and C18-UFLC method for selective quantification of selected chemical forms of tocopherol and tocotrienol in diverse foods. Food Chemistry, 437, 137909, https://doi.org/10.1016/j.foodchem.2023.137909; MNiSW 200 pkt. IF 8,8 Q1
- Czerwonka M., Białek A., Bobrowska-Korczak B. 2024. A novel method for the determination of squalene, cholesterol and their oxidation products in food of animal origin by GC-TOF/MS. International Journal of Molecular Sciences. 25 (5), 2807, https://doi.org/10.3390/ijms25052807
MNiSW 140 pkt. IF 4,9 Q1 - Czerwonka M., Białek A., Skrajnowska D., Bobrowska-Korczak B. 2024. Evaluation and discrimination of lipid components and iron and zinc levels in chicken and quail eggs available on the Polish market. Foods. 13, 10, 1571, https://doi.org/10.3390/foods13101571 MNiSW 100 IF 4,7 Q1
- Czerwonka M., Gielecińska A., Białek A., Białek M., Bobrowska-Korczak B., 2024. Cholesterol and Its oxidation derivatives content in market dairy products. Nutrients 16, 1371, https://doi.org/10.3390/nu16091371 MNiSW 140 IF 4,8 Q1
- Ferenc K., Marcinkowski M., Olszewski J., Kowalczyk P., Pilžys T., Garbicz D., Dib N., Świderska B., Matyba P., Gajewski Z., Grzesiuk E., Zabielski R. 2024. The proteomic profile is altered but not repaired after bariatric surgery in type 2 diabetes pigs. Scientific Reports 14, 10235. https://doi.org/10.1038/s41598-024-60022-9 MNiSW 140 pkt.. IF 3,8 Q1
- Głogowski R., Clauss M., Bełżecki G. 2024. Biochemical properties and hydrolytic activity in the nutria (Myocastor coypus) digestive tract. Animal Science and Genetics 20, 1, 3-10. https://doi.org/10.5604/01.3001.0054.3912 MNiSW 70 pkt; IF= 1,8; Q3
- Jelińska M., Białek A., Czerwonka M., Skrajnowska D., Stawarska A., Bobrowska-Korczak B. 2024. Knowledge of Food–Drug Interactions among Medical University Students. Nutrients 16, 15, 2425, https://doi.org/10.3390/nu16152425 MNiSW 140 pkt. IF 4,8 Q1
- Jęczmyk A., Krych S., Jekiełek M., Jurkiewicz M., Kowalczyk P., Kramkowski K., Hrapkowicz T. 2024. Wound healing complications after sternotomy-causes, prevention, and treatment-a new look at an old problem. Journal of Clinical Medicine. 13, 7431, https://doi.org/10.3390/jcm13237431
MNiSW 140 pkt. IF 3,0 Q1 - Johne M., Maculewicz E., Mastalerz A., Białek M., Wojtak W., Osuch B., Majewska M., Czauderna M., Białek A. 2024. Dietary Patterns, Serum BDNF and Fatty Acid Profiles in Physically Active Male Young Adults: A Cluster Analysis Study. Nutrients. 16(24), 4326, https://doi.org/10.3390/nu16244326
MNiSW 140 pkt. IF 4,8 Q1 - Konieczka P., Przybylski W., Jaworska D., Żelechowska E., Sałek .P, Szkopek D., Drażbo A., Kozłowski K., Jankowski J. 2024. Myofibrillar protein profile of the breast muscle in turkeys as a response to the variable ratio of limiting amino acids in feed. Agriculture 14, 2, 197, https://doi.org/10.3390/agriculture14020197 MNiSW 100 pkt. IF 3,3 Q1
- Konopka A., Gawin K., Barszcz M. 2024. Hedgehog signalling pathway and its role in shaping the architecture of intestinal epithelium. International Journal of Molecular Sciences. 25, 12007, https://doi.org/10.3390/ijms252212007 MNiSW 140 pkt. IF 4,9 Q1
- Kowalczyk P., Krych S., Kramkowski K., Jęczmyk A., Hrapkowicz T. 2024. Effect of oxidative stress on mitochondrial damage and repair in heart disease and ischemic events. International Journal of Molecular Sciences. 25, 12467, https://doi.org/10.3390/ijms252212467 MNiSW 140 pkt. IF 5,6 Q1
- Krych S., Jęczmyk A., Jurkiewicz M., Żurek M., Jekiełek M., Kowalczyk P., Kramkowski K., Hrapkowicz T. 2024. Viral Myocarditis as a Factor Leading to the Development of Heart Failure Symptoms, Including the Role of Parvovirus B19 Infection—Systematic Review. International Journal of Molecular Sciences 25, 8127, https://doi.org/10.3390/ijms25158127 MNiSW 140 pkt. IF 5,6 Q1
- Lauková A., Chrastinová Ľ., Focková V., Plachá I., Bino E., Grešáková Ľ., Formelová Z., Žitňan R., Belzecki G., Miltko R., Pogány Simonová M. 2024. Assessment of postbiotic, mundticin-like substance EM 41/3 application in broiler rabbits. Applied Sciences 14, 12, 5059, https://doi.org/10.3390/app14125059 MNiSW 100 pkt. IF 2,5 Q3
- Lauková A., Chrastinová L., Plachá I., Focková V., Zábolyová N., Bino E., Grešáková L., Žitňan R., Formelová Z., Ščerbová J., Belzecki G., Miltko R., Pogány Simonová M. 2024. Dairy-derived and bacteriocin-producing strain Lactiplantibacillus plantarum LP17L/1: An assessment of Its safety and effect using broiler rabbits. Frontiers in Bioscience – Elite16, 3, 21, https://doi.org/10.31083/j.fbe1603021 MNiSW 70 pkt. IF 0
- Lepionka T., Białek M., Czauderna M., Wojtak W., Maculewicz E., Białek A. 2024. Exploring the influence of the selected conjugated fatty acids isomers and cancerous process on the fatty acids profile of spleen. Cancers 16, 3, 479, https://doi.org/10.3390/cancers16030479 MNiSW 140 pkt. IF 4,5 Q1
- Majewska M.P., Wolska-Świętlicka U., Miltko R., Bełżecki G., Kędzierska A., Kowalik B. 2024. The effect of natural and synthetic zeolites on polysaccharidase activity in the rumen of Jersey heifers. Journal of Animal and Feed Sciences 33, 4, 535-541, https://doi.org/10.22358/jafs/194302/2024 MNiSW 100 pkt. IF 1,4 Q3
- Miltko R., Majewska M.P., Frączek N., Kowalik B., 2024. Evaluation of the biochemical composition of the greater duckweed Spirodela polyrhiza
(L. Schleiden). Journal of Animal and Feed Sciences 33, 4, 542-546, https://doi.org/10.22358/jafs/193033/2024 MNiSW 100 pkt. IF 1,4 Q3 - Miltko R., Majewska M.P., Wojtak W., Białek M., Kowalik B., Czauderna M. 2024. Comparing the chemical composition of lesser duckweed (Lemna minor L.) grown in natural and laboratory settings. Journal of Animal and Feed Sciences 33, 3, 357-367, https://doi.org/10.22358/jafs/189963/2024
MNiSW 100 pkt. IF 1,4 Q3 - Paslawski R., Kowalczyk P., Paslawska U., Wiśniewski J., Dzięgiel P., Janiszewski A., Kiczak L., Zacharski M., Gawdzik B., Kramkowski K., Szuba A. 2024. Analysis of the model of atherosclerosis formation in pig hearts as a result of impaired activity of DNA repair enzymes. International Journal of Molecular Sciences 25, 4, 2282, https://doi.org/10.3390/ijms25042282 MNiSW 140 pkt. IF 5,6 Q1
- Sobol M., Skiba G., Kowalczyk P., Świątkiewicz M., Grela E.R. 2024. Markers of bone turnover and biomechanical properties of the third metacarpal bone of growing pigs subjectet to the different dietary phosphorus and calcium content. Annals of Animal Science 24, 2, 479–490 https://doi.org/10.2478/aoas-2024-0010 140 pkt. IF 1,8 Q2
- Szkopek D., Mendel M., Kinsner M., Fotschki B., Juśkiewicz J., Kozłowski K., Matusevičius P., Konieczka P. 2024. Interaction Between Peroxisome Proliferator-Activated Receptors and Cannabidiol in the Gut of Chickens Applied to Different Challenge Conditions. International Journal of Molecular Sciences25, 21, 11398, https://doi.org/10.3390/ijms252111398 MNiSW 140 pkt 4,9 Q1
- Szkopek D., Pierzynowski S.G., Pierzynowska K., Zaworski K., Kondej A Pancreatic., Wychowański P., Konieczka P., Seklecka B., Donaldson J., Jank M., Woliński J. 2024. A review: enzymes in the treatment of chronic pancreatic insufficiency in companion animals. Journal of Veterinary Internal Medicine 38, 2, 2026-2033, https://doi.org/10.1111/jvim.17096 MNiSW 200 pkt. IF 2,1 Q1
- Świątkiewicz M., Szczepanik K., Gala Ł., Grela E.R., Witaszek K., Barszcz M., Tuśnio A., Taciak M. 2024. Determination of the impact of extruded soybean press cake on rearing and health indices of piglets. Agriculture. 14, 1899, https://doi.org/10.3390/agriculture14111899 MNiSW 100 pkt. IF 3,3 Q1
- Święch E. 2024. Effect of dietary wheat gluten levels on intestinal mucin flow and composition in young pigs.Journal of Animal and Feed Sciences33, 517-524, https://doi.org/10.22358/jafs/189648/2024 MNiSW 100 pkt. IF 1,4 Q3
- Wavhal D.S., Koszelewski , Gulko C., Kowalczyk P., Brodzka A., Kramkowski K., Ostaszewski R. 2024. Mystery of the Passerini Reaction for the Synthesis of the Antimicrobial Peptidomimetics against Nosocomial Pathogenic Bacteria. International Journal of Molecular Sciences25, 15, 8330, https://doi.org/10.3390/ijms25158330 MNiSW 140 pkt. IF 4,9 Q1
- Wojtak W., Białek M., Białek A., Molik E., Czauderna M. 2024. Content of orotic acid and selected bioactive compounds in ovine milk during lamb rearing. Journal of Animal and Feed Sciences 33, 547-561, https://doi.org/10.22358/jafs/188489/2024 MNiSW 100 pkt. IF 1,4 Q3
- Wójcik M., Zieba D.A., Bochenek J., Krawczyńska A., Barszcz M., Gajewska A., Antushevich H., Herman A.P. 2024. The Effect of Endotoxin-Induced Inflammation on the Activity of the Somatotropic Axis in Sheep. International Journal of Inflammation 1057299, https://doi.org/10.1155/2024/1057299 MNiSW 140 pkt. IF 2,6 Q3
- Wypych A., Lepczyński A., Ożgo M., Bernaciak M., Herosimczyk A., Barszcz M., Gawin K., Ciechanowicz A.K., Kucia M., Pierzchała M. 2024. Effect of feeding high fat diets differing in fatty acid composition on oxidative stress markers and protein expression in mouse kidney. Journal of Animal and Feed Sciences 33, 2, 170-184, https://doi.org/10.22358/jafs/175920/2024 MNiSW 100 pkt. IF 1,4 Q3
Laboratorium Analizy Bariery Ochronnej Przewodu Pokarmowego


Kierownik Laboratorium
dr inż. Marcin Barszcz
🖂 m.barszcz@ifzz.pl
Laboratorium Analizy Bariery Ochronnej Przewodu Pokarmowego jest nową jednostką organizacyjną Zakładu Żywienia Zwierząt utworzoną w 2018 roku dzięki funduszom przyznanym przez Ministerstwo Nauki i Szkolnictwa Wyższego w ramach wniosku na finansowanie kosztów restrukturyzacji Instytutu w latach 2018-2019.
Tematyka badawcza
- Laboratorium prowadzi badania nad układem odpornościowym przewodu pokarmowego zwierząt gospodarskich i laboratoryjnych.
- Celem badań jest określenie wpływu żywienia (diety, składników pokarmowych, związków bioaktywnych) na odporność wrodzoną
i nabytą przewodu pokarmowego, a także roli mikrobioty jelitowej w kształtowaniu funkcji barierowej. - Laboratorium wykonuje analizy materiału biologicznego (tkanki, płyny ustrojowe i in.) na zlecenie i jest otwarte na współpracę z krajowymi
i zagranicznymi jednostkami naukowymi oraz innymi podmiotami.
Zakres usług oferowanych przez Laboratorium
- Laboratorium dysponuje szerokim zakresem metod analitycznych, umożliwiających oznaczanie parametrów specyficznych dla bariery ochronnej przewodu pokarmowego i funkcjonowania układu odpornościowego.
- Posiadana aparatura umożliwia również wykonywanie analiz niezwiązanych ze specyfiką Laboratorium.
- W celu przygotowania materiału biologicznego do badań oraz w większości analiz wykonywanych w Laboratorium wykorzystywana jest również
inna aparatura naukowo-badawcza Zakładu Żywienia Zwierząt.
Rodzaj próbek
- tkanki zwierzęce, homogenaty, supernatanty
- zawiesiny komórek
- płyny ustrojowe
- w zależności od analizy do Laboratorium mogą być przyjmowane próbki świeże lub mrożone.
Analizy
Cytometr przepływowy BD FACSCelesta:
- subpopulacje komórek układu odpornościowego (izolowane z przewodu pokarmowego i krwi)
- funkcje granulocytów i monocytów:
− fagocytoza
− wybuch
− tlenowy inne
Spektrofluorymetr SpectraMax iD3
- stężenie mucyn
- stężenie wydzielniczej immunoglobuliny A oraz innych przeciwciał i białek (ELISA)
- aktywność enzymów antyoksydacyjnych
- inne analizy mikropłytkowe oparte na pomiarach absorbancji, fluorescencji i luminescencji
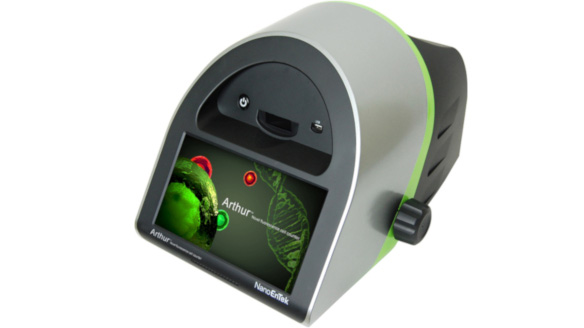
Licznik komórek Arthur:
- liczba komórek w zawiesinie (standaryzacja do cytometrii przepływowej i innych analiz)
- żywotność komórek (metoda z jodkiem propidyny)
- cykl komórkowy (metoda z jodkiem propidyny)
- apoptoza (metoda z aneksyną V i jodkiem propidyny)
- ekspresja białka zielonej fluorescencji (GFP)
Termocykler MIC:
- ekspresja genów kodujących m.in. mucyny, cytokiny, białka transportujące, microRNA
Analiza histologiczna
- obróbka chemiczna i zatapianie materiału biologicznego w bloczkach parafinowych
- cięcie materiału biologicznego zatopionego w bloczkach parafinowych (mikrotom rotacyjny HM355S)
- barwienie preparatów histologicznych:
− hematoksylina-eozyna (HE)
− floksyna-tartrazyna (FT)
− alcjan błękitu-kwas nadjodowy-odczynnik Schiffa (AB-PAS)
− błękit toluidyny (TB)
Analiza preparatów pod mikroskopem:
- liczba limfocytów śródnabłonkowych w jelicie cienkim i grubym świń (HE)
- liczba komórek tucznych (TB)
- liczba i rodzaj komórek kubkowych (AB-PAS)
- liczba komórek Panetha w jelicie cienkim gryzoni laboratoryjnych (FT).
Oferta Laboratorium wkrótce zostanie rozszerzona o inne metody analityczne, które są aktualnie opracowywane i wdrażane.
Wyposażenie

ANALIZATOR SEAHORSE XF Pro
Analizator metabolizmu komórkowego w czasie rzeczywistym Seahorse XF Pro (Agilent Technologies, Inc., Lexington, MA, USA ) umożliwia automatyczny, równoczesny pomiar dwóch głównych szlaków metabolicznych odpowiedzialnych za produkcję energii w komórkach: glikolizy i oddychania mitochondrialnego. Pomiary tempa zużycia tlenu i stopnia zakwaszenia pozakomórkowego prowadzone są za pomocą sensorów tlenu i pH w czasie rzeczywistym, przyżyciowo, w komorach w układzie płytek 96-dołkowych. Seahorse XF Pro umożliwia przeprowadzanie kompleksowych badań szlaków metabolicznych, procesów katabolicznych i anabolicznych oraz analizy wykorzystania surowców energetycznych przez nienaruszone i w pełni funkcjonalne komórki. Aparatura jest przystosowana do badania szerokiego spektrum komórek ssaków, a także komórek drożdży, wyizolowanych mitochondriów, całych organizmów wielokomórkowych, np. Caenorhabditis elegans oraz tkanek roślinnych. Dzięki portom iniekcyjnym analizator może dozować do czterech badanych substancji chemicznych do każdej z komór pomiarowych i prowadzi pomiary parametrów metabolicznych przed lub po iniekcji danej substancji.
CYTOMETR PRZEPŁYWOWY BD FACSCelesta
Cyfrowy cytometr przepływowy BD FACSCelesta (Becton Dickinson, Franklin Lakes, New Jersey, USA) umożliwiający jednoczesny pomiar 12 fluorescencji na jednej komórce, wyposażony w 3 lasery (niebieski, czerwony, fioletowy), podajnik próbek z płytek wielodołkowych oraz moduł FACS Flow Supply System do podawania płynu roboczego ze zbiornika 20 l i odprowadzania ścieków. Stosowany w badaniach z zakresu immunologii (m.in. populacje komórek, testy fagocytozy, wybuch tlenowy), biologii komórki (m.in. apoptoza, cykl komórkowy) i in.


SPEKTROFLUORYMETR SPECTRAMAX ID3
Wielofunkcyjny czytnik płytek SpectraMax iD3 (Molecular Devices, San Jose, California, USA) umożliwiający odczyty absorbancji, fluorescencji (z góry i z dołu), luminescencji oraz pomiary kinetyczne. Posiada termostatowaną komorę pomiarową z możliwością regulacji temperatury (od 5°C powyżej temperatury otoczenia do 66°C) oraz wytrząsania płytki. Wykorzystywany do analiz biochemicznych, testów immuno-enzymatycznych (ELISA) i in.
AUTOMATYCZNY LICZNIK KOMÓREK ARTHUR
Automatyczny, fluorescencyjny licznik komórek Arthur (NanoEnTek, Seul, Korea Południowa) to 3-kanałowy licznik (jasne pole, zielona fluorescencja, czerwona fluorescencja) umożliwiający szybką analizę liczby komórek w zawiesinie, ekspresji białek zielonej i czerwonej fluorescencji, żywotności komórek, apoptozy i cyklu komórkowego w próbach o objętości 25 µl. Licznik może być używany do różnego typu komórek eukariotycznych.

TERMOCYKLER MIC DO REAL-TIME PCR
Termocykler MIC (Bio Molecular Systems, Upper Coomera, Australia) do analizy ekspresji genów metodą real-time PCR, oparty na technologii indukcji magnetycznej, model 4-kanałowy, z systemem rotorowym na 48 prób zapewniającym stałą drogę optyczną, jednorodność temperatury i dokładność. Umożliwia przeprowadzenie PCR w objętości reakcyjnej od 10 do 25 µl.
INKUBATOR CO2 MIDI 40
Inkubator CO2 MIDI 40 (Thermo Scientific, Waltham, Massachusetts, USA) do hodowli komórkowych zapewniający odpowiednie stężenie CO2, wilgotność powietrza i temperaturę, z płaszczem powietrznym i komorą roboczą ze stali nierdzewnej o pojemności 40 l. Inkubator przeznaczony do krótko-terminowych hodowli limfocytów (testy proliferacji).


AUTOMATYCZNY MIKROTOM ROTACYJNY HM355S
Automatyczny mikrotom rotacyjny HM355S (Thermo Shandon Limited, Runcorn, UK) z systemem transferu skrawków (STS) po torze wodnym i przystawką chłodzącą Cool-Cut do precyzyjnego cięcia materiału biologicznego zatopionego w bloczkach parafinowych. Mikrotom umożliwia uzyska-nie preparatów histologicznych o grubości od 0,5 do 100 µm. System Cool-Cut utrzymuje przez długi czas niską tempera-turę bloczku parafinowego w uchwycie i wraz z STS zapewnia maksymalną elastyczność skrawków.
SUSZARKA FN500
Suszarka FN500 (Nüve, Ankara, Turkey) o pojemności użytkowej 110 l, z wymuszonym obiegiem powietrza zapewniającym wysoką jednorodność i stabilność temperatury w komorze, sterowana mikroprocesorowo. Zakres temperatury: od +5°C powyżej temperatury otoczenia do 250°C. Wykorzystywana do suchej sterylizacji narzędzi, suszenia preparatów histologicznych oraz inkubacji prób w czasie analiz.

Kontakt:
Instytut Fizjologii i Żywienia Zwierząt im. Jana Kielanowskiego Polskiej Akademii Nauk
ul. Instytucka 3, 05-110 Jabłonna
dr inż. Marcin Barszcz
🖂 m.barszcz@ifzz.pl; tel. 22 765 33 00; 22 765 33 30